Fossilien in den Höhlenlehmen der Barbarossahöhle, Kyffhäuser von ANJA ADLER & DOROTHEE MERTMANN Zusammenfassung Abstract Resumé
Abb. 1: Die Barbarossahöhle im Geopark Kyffhäuser, Weg vom Eingangsbereich zur Neptungrotte
2. Geographischer Überblick
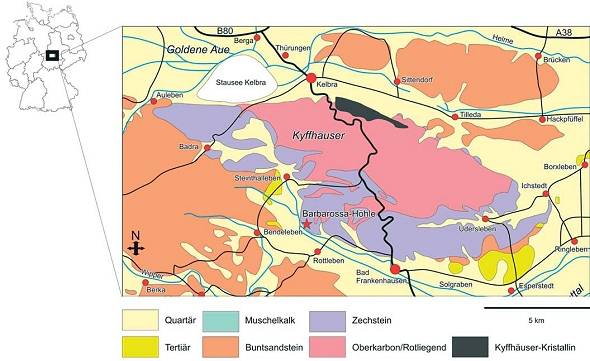
Abb. 2: Geologische Karte des Untersuchungsgebietes (nach GEOPARK KYFFHÄUSER 2005)
4. Höhlenlehme der Barbarossahöhle 4.1 Lokationen und Granulometrie Lokalität 2 Lokalität 3
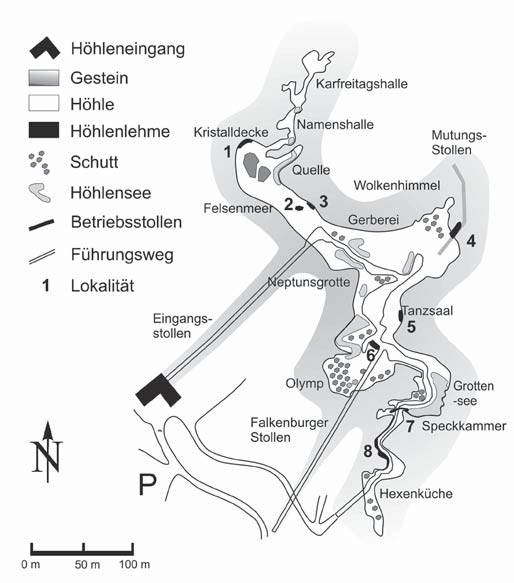
Abb. 3: Schematischer Grundriss der Barbarossahöhle mit Höhlenräumen und Karstgewässern (nach BRUST & HÖFER 2004) und untersuchten Höhlenlehmvorkommen 1 - 8Lokalität 4 Die vierte Lokalität befindet sich am Eingang zum Bergbaustollen (Abb. 6). Der Höhlenlehm weist eine Mächtigkeit von mindestens 1,50 m auf und kann über weite Teile des Stollens an der Decke des Stollens in dann reduzierter Mächtigkeit von ca. 40 cm verfolgt werden. Eine Schichtung ist deutlich durch Korngrößenunterschiede zwischen Silt und Feinkies sowie ocker bis rotbraune Farben angezeigt (Abb. 7). Flach einfallende, gebogene Schrägschichtung ist in einigen Schichten vorhanden. Damit liefert dieses Vorkommen aufgrund der Grobkörnigkeit der Schichten und der gefundenen Schrägschichtung einen Hinweis auf ein höher energetisches, fluviatiles System, das die Höhle durchströmt haben muss. Die Proben sind allerdings weitgehend fossilarm bis fossilfrei. Lokalität 5 Lokalität 6 Lokalität 7
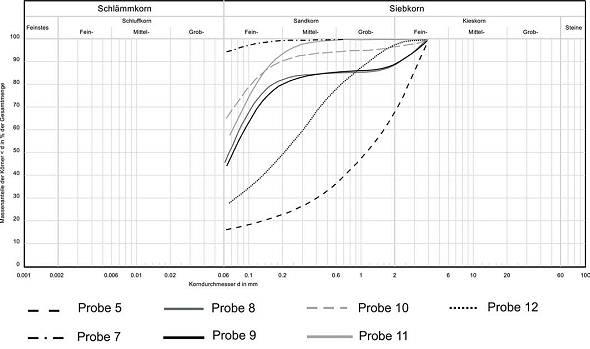
Abb. 4: Vorkommen der Höhlenlehme im Bereich der Kristalldecke an Lokalität 1 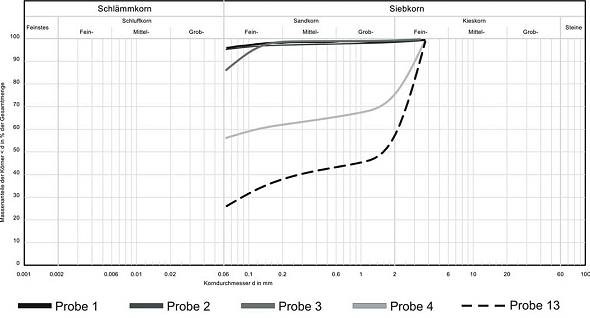 Abb. 5: Korngrößenverteilungen ausgewählter Proben: Lokalität 1 (Probe 1 + 2), Lokalität 2 (Probe 3), Lokalität 3 (Probe 4), Lokalität 5 (Probe 13)Lokalität 8 Aufschluss 8 befindet sich nahe am Höhlenausgang. Mit etwa 15 m Länge und einer Höhe von 1 m bis 1,50 m ist es zudem das größte ausstreichende Vorkommen. Bereiche mit und ohne Schichtung sowie unterschiedlicher Farbe werden unterschieden: Am nördlichen Ende des Aufschlusses zeigt sich eine deutliche Schichtung des Höhlenlehms: Der untere Bereich (30 cm) wird von einem dunklen, feingliedrig geschichteten Lehm, in dem zum Teil größere (> 5 cm) Gipsrosetten eingeschaltet sind (Abb. 9), dominiert. Ein bis zu 15 cm dicker harter Horizont aus Sulfat mit wenig Pelit grenzt den oberen, im cm-Bereich geschichteten Bereich, nach unten ab. Nach ca. 5 m zeigt sich ein deutlicher Farbwechsel der Höhlenlehme von rötlich zu hell- bis dunkelbraun. Gleichzeitig verschwindet die deutliche Schichtung. Weiter südlich geht die Farbe dann erneut in einen Rotton über, der den weiteren Verlauf des Vorkommens bestimmt. Sedimentologisch ergab sich bei allen Proben die für die Höhlenlehme typische Dominanz an Schlämmkorn (< 0,063 mm; 90 - 97 %). Gröbere Fragmente bestanden aus Sulfat. Gelegentlich ist Kohle eingestreut.
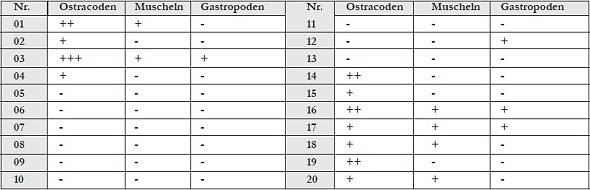
Abb. 6: Vorkommen der Höhlenlehme im Bereich des Kupferschieferstollens an Lokalität 44.2 Die Faunen der Höhlenlehme Die Aufbereitung der 20 Proben zeigte, dass nur in 10 Proben Fossilien enthalten sind (Abb. 10). Diese sind vor allem Ostrakoden, die in sehr unterschiedlichen Häufigkeiten auftreten, sowie meistens schon makroskopisch erkennbare Muscheln und Gastropoden. Letztere treten nie unter 1 mm Korngröße auf. Die Ostrakoden fanden sich meistens in der Kornfraktion größer 250 μm, nur in Ausnahmen darunter oder in der Kornfraktion größer 500 μm. Die Bestimmung der Ostrakoden erfolgte vor allem nach der äußeren Gestalt, den peripheren Porenkanälen, der Schalenoberfläche und der Innenlamelle. In den Proben sind selten komplette Gehäuse erhalten; zum größten Teil wurde nur eine Schalenhälfte zur Bestimmung herangezogen. Herr Prof. Dr. Steffen Mischke führte die Bestimmung der Ostrakoden maßgeblich durch.
Abb. 7: Korngrößenverteilungen der im Profil an Lokalität 4 entnommenen Proben
Ilyocypris bradyi
Abb. 8: Vorkommen der Höhlenlehme im Bereich des Grottensees an Lokalität 7  Abb. 9: In den Höhlenlehmen vorkommende Gipsrosetten, hier an Lokalität 8
Abb. 10: Die Faunen der Höhlenlehme der Barbarossahöhle, unterteilt in Ostrakoden, Muscheln und Gastropoden, und ihr Vorkommen in den entnommenen Proben: (+) geringe Häufigkeit; (++) häufiges Auftreten; (+++) sehr häufiges, dominantes Vorkommen; (-) keine Individuen enthaltenCandona candida und Candona neglecta Charakteristisch sind die eher dreieckige Form von Candona candida und die bohnenförmige Gestalt von Candona neglecta. Beide Schalen sind glatt. Die Gattung Candona kommt meist in stehenden Süßwasserhabitaten bei eher niedrigen Temperaturen vor. Höhere Salzgehalte im Grundwasser oder in Quellgewässern toleriert sie dagegen im Allgemeinen eher nicht. Candona candida besiedelt zum Beispiel auch Lockersedimente der Uferbereiche von Seen (MARMONIER 1984, 1985). Als besonderes Charakteristikum zeigen sich die Individuen tolerant gegenüber dem pH-Wert, auch wenn er im sauren Milieu liegt (HILLER 1972, HARTMANN & HILLER 1977). Nach HILLER (1972) toleriert sie einen maximalen Salzgehalt von 5,77 ‰. Sie sind weit verbreitet, zum Teil auch im polaren Bereich. Überliefert ist dieses Taxon seit dem Oligozän (HELMDACH 1977), Unteren Pliozän (MEISCH 2000) und vermehrt ab dem Pleistozän bis rezent. Candona neglecta kommt sowohl in kleinen Wasserläufen als auch in Teichen und Seen vor, bevorzugt dabei niedrigenergetische Milieus mit einer geringen Wassertiefe. Sowohl die larvalen als auch die adulten Stadien sind resistent gegenüber Austrocknung, sodass auch temporäre Gewässer besiedelt werden können. Die Art vermag hypoxische Bedingungen mit einem Sauerstoffgehalt unterhalb von 3 mg/l besonders im Sommer zu überleben (DANIELOPOL et al. 1985, 1993) und toleriert auch eine Salinität bis 16 ‰ und Ca-Gehalte von über 72 mg/l (MEISCH 2000). Bekannt ist diese Art seit dem Pleistozän bis rezent.
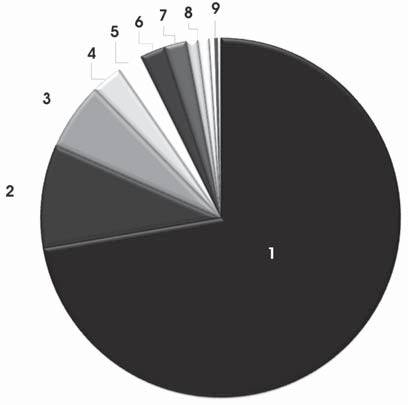
Abb. 11: Schematische Darstellung des prozentualen Auftretens der einzelnen Arten in der Probe 3: 1. Ilyocypris sp., 2. Candona neglecta, 3. Pseudocandona marchica, 4. Cyclocypris laevis, 5. Heterocypris incongruens, 6. Prionocypris zenkeri, 7. Heterocypris salina, 8. Candona candida, 9. Cyprideis torosa, Tonnacypris sp., Eucypris sp. und Potamocypris sp.Cyclocypris laevis Die Form von Cyclocypris laevis ist glatt und oval. Charakteristisch ist der Unterschied zwischen dem glatten ventralen und dem stark konvexen dorsalen Rand der Schalen. Ebenso gut erkennbar ist die Innenlamelle, vor allem im Bereich des Posteriors. Cyclocypris laevis kommt sowohl im permanenten als auch im temporären Süßwasser sowie in leicht salzigen Gewässern und Teichen bei niedriger und hoher Fließgeschwindigkeit vor. Sie bevorzugt dabei Gewässer mit erhöhten Calciumgehalten.
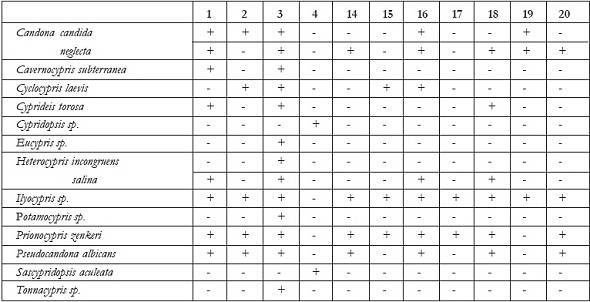
Abb. 12: Das Vorkommen der einzelnen Arten in den fossilführenden Proben: (+) kommt vor; (-) kommt nicht vorHeterocypris incongruens und Heterocypris salina Heterocypris weist im Gegensatz zu anderen Formen eine auffallend breite Innenlamelle auf. Anterior und Posterior sind weit gerundet und relativ breit. Die Form ist länglich mit einem schwach konkaven ventralen Rand und einem gewölbten dorsalen Rand. Dieser ist bei Heterocypris salina deutlich spitzer als bei Heterocypris incongruens. Letztere bevorzugt kleine, temporäre Wasserkörper mit tonigem Substrat und wenig Vegetation. Funde zeigen, dass sie vor allem in flachen Gewässern vorkommt. Vor allem in deren Randbereichen siedeln sich zuweilen große Populationen an. Zudem ist Heterocypris incongruens auch aus dem Grundwasser, aus Höhlen und sauerstoffarmen Habitaten bekannt (MEISCH 2000). Fossil ist Heterocypris incongruens seit dem Pleistozän überliefert. Heterocypris salina bevorzugt ebenso kleine, flache, eher temporäre aquatische Habitate. Der Salzgehalt der Gewässer sollte optimal etwa 6 ‰ betragen (GANNING 1967, 1971). Sie toleriert aber auch geringere oder höhere Salzgehalte, höhere Calciumgehalte und ein weites Temperaturspektrum (MEISCH 2000). Zudem kann sie angepasst an Dunkelheit und Vegetationsarmut existieren und ist damit auch typisch für Höhlengewässer. Die Art ist seit dem Miozän bekannt (MEISCH 2000). Pseudocandona albicans Cyprideis torosa Prionocypris zenkeri Cavernocypris subterranea Sarscypridopsis aculeata Tonnacypris sp. Potamocypris sp. Eucypris sp. Cypridopsis sp.
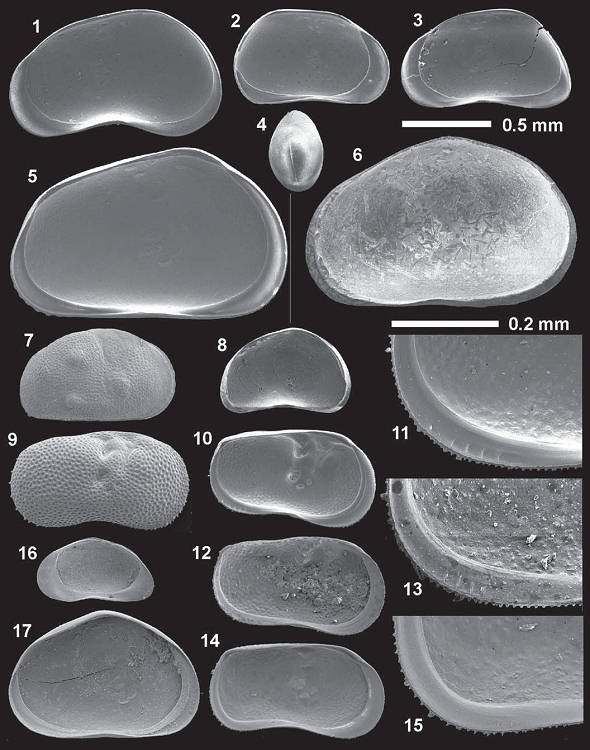
Abb. 13: Ostrakoden der Barbarossahöhle: 1. Candona neglecta, rechte Klappe (RV) Innenansicht (iv); 2. Pseudocandona albicans, linke Klappe (LV) iv; 3. Candona candida, RV iv; 4. Cyclocypris laevis, dorsal Schale; 5. Prionocypris zenkeri, LV iv; 6. Heterocypris incongruens, RV iv; 7. Cyprideis torosa forma torosa, RV Außenansicht (ev); 8. Potamocypris sp., LV iv; 9.-15. Ilyocypris bradyi, 9. RV ev; 10. LV iv, 11. Vergrößerung des posteroventralen Teils des unteren linken Abschnitts der Form, vier Rippen sind deutlich erkennbar; 12. LV iv, 13. Vergrößerung des posteroventralen Teils des unteren linken Abschnitts der Form, vergleichbar 11, 14. LV iv, 15. Vergrößerung des posteroventralen Teils des unteren linken Abschnitts der Form, zwei randliche Rippen sind schwach sichtbar; 16. Cavernocypris subterranea, RV iv; 17. Heterocypris salina, LV iv.Weiterhin weisen fast alle Arten eine glatte Oberfläche mit einer breiten Innenlamelle auf. Eine Artzuordnung konnte allerdings nicht durchgeführt werden. Die Formen leben in den verschiedensten nicht-marinen, zumeist permanenten, vegetationsreichen Habitaten, die nicht direkt der Sonne ausgesetzt sind, zum Teil auch in Quellen oder wie rezente Vertreter, in Wasserreservoiren (MEISCH 2000). Fossile Funde stammen vor allem aus dem Pleistozän bis rezent. 4.2.2 Gastropoden und Pelecypoden
Abb. 14: Mollusken der Barbarossahöhle: 1. Vallona sp., Umbilikal; 2. Vertigo sp., 3. - 4. Pisidium subtruncatum (3. linke Schale, 4. Vergrößerung des Scharniers)5. Schlussfolgerungen Die Höhlenlehme zeigen überwiegend eine typische feinkörnige Konsistenz fast ausschließlich aus Schlämmkorn kleiner < 0,063 mm, die sich in den Kornsummenkurven darstellt. Nur in Lokalität 4 lässt sich ein deutlich erhöhter Gehalt an gröber körnigen, vor allem sandigen Bestandteilen ermitteln, der sich in schräggeschichteten Lagen konzentriert. Der bei den anderen Lokalitäten festgestellte Anteil an Grobkorn besteht aus Anhydrit und Gips, entstammt also dem Nebengestein. Nach der Form der Höhlenlehmvorkommen und ihrem internen Gefüge unterscheiden sich drei unterschiedliche Typen. Anschnitte von ehemaligen Zuflusskanälen sind durch eine gerade Basis und eine nach oben gewölbte Form charakterisiert (z.B. Lokalität 3). Hier könnte Wasser in die Höhle eingedrungen sein und mitgebrachtes Sediment plombiert den Zuflusskanal. Rissfüllungen (z.B. in den Aufschlüssen 5 und 7) sind ungeschichtet, schmal und keilen lateral aus. Sie sind an Risse im Nebengestein, ob als echte Klüfte oder lösungsbedingte Spalten kann nicht entschieden werden, gebunden. Einige ausgedehntere Vorkommen an Höhlenlehmen sind deutlich geschichtet. Hier schließen wir auf ein unterirdisches Gewässersystem mit unterschiedlichen Fließgeschwindigkeiten (Lokalität 1, 4, 6 und 8). Meist handelt es sich um eher niedrigenergetische Systeme vergleichbar den heutigen Seen in der Höhle. Nur in Lokalität 4 zeigen die schräggeschichteten groben Lagen einen zwischenzeitlich hochenergetischen Transport an. Eine Einordnung von Lokalität 2 in die genannten Typen kann aufgrund der geringen Größe bzw. Zugänglichkeit und der fehlenden Gefüge nicht vorgenommen werden. Über das Entstehungsalter der Höhlenlehme und die Zeitlichkeit der Höhlenentstehung lassen sich nur wenige Aussagen treffen. Einige Ostrakodenformen sind bereits seit dem Oligozän, Miozän beziehungsweise dem Pliozän bekannt. Die meisten Formen kommen allerdings erst seit dem Pleistozän vor und sind auch noch rezent bekannt. Somit lässt der Fossilinhalt der Höhlenlehme direkt keine bessere Eingrenzung als Quartär zu. Auch die Kohlestückchen verheißen dazu wohl keine weiteren Erkenntnisse, da sie wahrscheinlich aus vormals auch oberirdisch anstehenden, der Höhle benachbarten Flözvorkommen der permokarbonen Molassefolgen stammen. Allerdings beschrieben FUHRMANN et al. (1990) aus dem Eem-Interglazial von Gröbern und Grabschütz Faunenvergesellschaftungen, die im Wesentlichen nun auch in den Höhlenlehmen gefunden wurden. Im Folgenden sollen die vorgefundenen Ostrakodenformen in Hinblick auf ihre ökologischen Ansprüche analysiert werden. Es wurden 15 Gattungen gefunden: Ilyocypris dominiert, gefolgt von Candona und Pseudocandona. Alle anderen Formen sind selten. Die bestimmten Ostrakodengattungen und -arten besitzen durchaus unterschiedliche ökologische Ansprüche. Zu klären ist die Frage, ob die Formen alle in der Höhle selbst lebten oder auch eingeschwemmte Formen vorhanden sind. Durchgeführte Messungen der heutigen Bedingungen in den Gewässern der Höhle zeigen ein Temperaturspektrum von 8,8 bis maximal 9,6 °C, sowie pH-Werte, die im Dezimalbereich um 6,7 schwanken. SPANGENBERG (1973) publizierte Messwerte zwischen 7,5 - 8,8 °C und damit niedrigere Temperaturwerte. Er bestätigte aber die nahezu neutralen pH-Bedingungen. Vegetation ist am Rande und in den Höhlengewässern nicht zu beobachten und auch nicht überliefert. SPANGENBERG (1973) beschrieb aus dem Umfeld der Höhle mehrere weitere Ostrakodenformen: Candona angusta, Candona husmanni, Candona vavrai, Cyclocypris ovum, Cyclocypris serena und Ilyodromus olivaceus. Im Höhlengewässer selbst fand er Candona sp. In Bezug auf die Temperatur zeigen die meisten Arten eine weit reichende Toleranz, wobei die meisten Formen kältere Gewässer bevorzugen. Aber auch Temperaturanstiege werden für eine geringe Zeitspanne geduldet. Es gibt reine Süßwasserformen (Tonnacypris, Potamocypris) neben Formen, die in leicht brackischen Gewässern ihr Optimum besitzen (Cyprideis, Heterocypris, Sarscypridopsis). Die Gattung Ilyocypris lässt sich auch in leicht salzigen Habitaten finden. Einige Arten, wie Pseudocandona albicans, Cyclocypris laevis, Heterocypris salina, Cyprideis torosa und Sarscypridopsis aculeata, weisen eine größere Toleranz gegenüber dem Salzgehalt auf. Auch aus dem Eem-Interglazial von Grabschütz wird die Kombination von Heterocypris salina, Candona angulata und Cyprideis torosa als Indikator der Versalzung eines aquatisch-glazialen Systems herangezogen (FUHRMANN et al. 1990). Cyprideis torosa, Heterocypris incongruens, Heterocypris salina sowie Pseudocandona marchica bevorzugen weitestgehend vegetationsarme Gewässer. Hingegen kommen Cypridopsis sp., Eucypris sp., Tonnacypris sp. und Prionocypris zenkeri fast ausschließlich in Gewässern mit einer üppigen Vegetation sowohl im Uferbereich als auch in größeren Wassertiefen vor. Die dominierende Art Ilyocypris bradyi toleriert beide Milieus. Pseudocandona marchica, Heterocypris incongruens und Heterocypris salina kommen allerdings eher in oberflächenunabhängigen Bereichen vor. Die gefundene Faunenvergesellschaftung weist ein weit reichendes Spektrum in Bezug auf die Art der aquatischen Systeme auf. Der größte Teil, allen voran Ilyocypris tendiert zu niedrig energetischen, z.T. stagnierenden Gewässern. Eucypris sp. und Sarscypridopsis aculeata bevorzugen dagegen deutlich höher energetische, fluviatile Systeme. Dahingegen sind Cyclocypris laevis, Cyprideis torosa, Heterocypris incongruens und -salina sowie Tonnacypris sp. in Bezug auf die Wasserenergie des Systems anpassungsfähig. Auch bei Betrachtung der Wasserführung der aquatischen Systeme kann keine einheitliche Aussage getroffen werden. Allgemein tolerieren die meisten Formen sowohl temporäre als auch permanente Gewässer. Alle Arten tolerieren auch die durch das anhydritische Nebengestein verursachten erhöhten Gehalte an Ca-und SO4- Ionen im Wasser. Wenn das heutige Gewässersystem ähnlich auch früher während der Bildungsphase der Höhlenlehme existiert hätte, müsste es überwiegend niedrigenergetisch, flach, ohne Vegetation und Lichteinstrahlung, limnisch bis gering brackisch und mit einem hohen Ca-Gehalt beschrieben werden. Ein großer Teil der beschriebenen Formen könnte also durchaus in der Höhle selbst beheimatet gewesen sein. Dies gilt vor allem für Heterocypris incongruens, Heterocypris salina, Candona neglecta, Sarscypridopsis aculeata sowie Pseudocandona marchica. Bei der dominierenden Ilyocypris bradyi kann die Besiedlung von Höhlengewässern nicht ausgeschlossen werden. Nur Cypridopsis sp., Eucypris sp., Tonnacypris sp. und Prionocypris zenkeri erscheinen problematisch, da sie zum Beispiel vegetationsreiche Standorte besiedeln. Sie scheinen also eher von außen zugeführt worden zu sein. Die erstmalige Untersuchung der Höhlenlehme ergibt also neben sedimentologischen Aspekten auch paläontologische Hinweise auf ein Leben in der Höhle. Dass allerdings alle bestimmten Formen dort beheimatet waren, kann nicht gefolgert werden. Vielmehr gehen wir davon aus, dass variable Zuflüsse von außen Material und Organismen einschwemmten. Ein Rezentvergleich zur Beziehung Pfannspring – Höhle könnte helfen, zusätzliche Informationen zu gewinnen. 6. Dank Literatur BRUST, M.K. (2005): Die Erschließung und Entwicklung der Barbarossahöhle als Schauhöhle. – Geowiss. Exkf. u. Mitt. DGG 225: 18 - 23 BRUST, M.K. (2008): Die „Mansfeldischen Kalkschlotten“ und ihre Bedeutung für den historischen Kupferschiefer-Bergbau. – EDGG Exkf. u. Veröff. Dt. Ges. f. Geowiss. 235: 10 - 18 BRUST, M.K. & HÖFER, M. (2004): Höhlenplan der Barbarossahöhle. http://www.hoehle.de DANIELOPOL, D.L., HANDL, M. & YIN, Y. (1993): Benthic ostracods in the pre-alpine deep lake Mondsee: Notes on the origin and distribution. – In: McKenzie, K.G. & Jones, P.J., Hrsg.: Ostracoda in the Earth and Life Sciences. Proceeding of the 11th International Symposium on Ostracoda, Warmambool, Voctoria, Australia, S. 465 - 480 DANIELOPOL, D.L., GEIGER, W., TÖLDERER-FARMER, M., ORELLANA, C.P. & TERRAT, M.N. (1985): The Ostracoda of Mondsee: spatial and temporal changes during the last 50 years. – In: Danielopol, D.L., Schmidt, R. & Schultze, E., Hrsg.: Contributions of the Paleolimnology of the Trumer Lakes (Salzburg) and the Lakes Mondsee, Attersee and Traunsee (Upper Austria). Limnologisches Institut der Österreichischen Akademie der Wissenschaften, Mondsee, S. 99 - 121 DECKER, M. (2009): Sanderz und Kupferschiefer in der Barbarossahöhle, Kyffhäuser. – Bachelorarbeit Institut für Geowissenschaften, Martin Luther-Universität Halle-Wittenberg, 39 S., unveröff. DECKER, M., MERTMANN, D. & FISCHER, H. J. (2011): Gesteine in der Barbarossahöhle, Kyffhäuser (Thüringen). – Aufschluss 62: 13 - 25 FRANZKE, H.-J., ZEH, A. & MEIER, S. (2007) Die metamorph-magmatische und strukturelle Entwicklung des Kyffhäuser Kristallins/ Mitteldeutsche Kristallinzone - Vergleich mit der Wippra-Zone und dem Eckergneis/Harz. – Z. geol. Wiss. 35 (1/2): 27 - 61 FUHRMANN, R. & PIETRZENIUK, E. (1990a): Die Ostracodenfauna des Interglazials von Gröbern (Kreis Gräfenhainichen). – In: Eissmann, L., Hrsg.: Die Eemzeit und die frühe Weiseleiszeit im Saale-Elbe-Gebiet: Geologie, Paläontologie, Paläökologie. Altenburger naturwiss. Forsch. 5: 168 - 193 FUHRMANN, R. & PIETRZENIUK, E. (1990b): Die Ostracodenfauna des Interglazials von Grabschütz (Kreis Delitzsch). – In: Eissmann, L., Hrsg.: Die Eemzeit und die frühe Weiseleiszeit im Saale-Elbe- Gebiet: Geologie, Paläontologie, Paläökologie. Altenburger naturwiss. Forsch. 5: 202 - 227 GANNING, B. (1967): Laboratory experiments in the ecological work on rockpool animals with special notes on the ostracoda Heterocypris salinus. – Helgoländer wiss. Meeresunt. 15: 27 - 40 GANNING, B. (1971): On the ecology of Heterocypris salinus, H. incongruens and Cypridopsis aculeata (Crustacea, Ostracoda) from Baltic brackish-water rockpools. – Marine Biology 8(4): 271 - 279 GEOPARK KYFFHÄUSER (2005): Geopfade – Unerwartete Begegnungen auf steinigen Wegen. – 128 S. HARTMANN, G. & HILLER, D. (1977): Beitrag zur Kenntnis der Ostracodenfauna des Harzes und seines nördlichen Vorlandes (unter besonderer Berücksichtigung des Männchens von Candona candida. – 125 Jahre Naturwissenschaftlicher Verein Goslar, S. 99 - 116 HELMDACH, F.-F. (1977): Leitfaden zur Bestimmung fossiler und rezenter Ostrakoden. – 264 S. HILLER, D. (1972): Untersuchungen zur Biologie und zur Ökologie limnischer Ostracoden aus der Umgebung von Hamburg. – Archiv f. Hydrobiologie, Suppl. 40 (4): 400 - 497 HOPF, H. (2000) Geotope des Kyffhäusers als Beispiel einer komplexen geoökologischen Einheit. – Geowiss. Mitt. Thüringen, Beih. 10: 137 - 149 KEMPE, S. (2008): Gipskarst – Ein Überblick. – EDGG Exkf. u. Veröff. Dt. Ges. f. Geowiss. 235: 30 - 41 KUPETZ, M. (2005): Gipskarst am Kyffhäuser und die Genese der Barbarossahöhle. – Geowiss. Exkf. u. Mitt. DGG 225: 12 - 17 KUPETZ, M. (2008): Neue Vorstellungen zur Genese von Höhlen vom Typ der „Mansfeldischen Kalkschlotten“. – EDGG Exkf. u. Veröff. Dt. Ges. f. Geowiss. 235: 19 - 29 MARMONIER, P. (1984): Vertical distribution and temporal evolution of the ostracod assemblage of the Seebach sediments (Lunz-Austria). – Jahresber. Biol. Stat. Lunz d. österr. Akad. d. Wiss. 7: 49 - 82 MARMONIER, P. (1985): Répartition spatial des Ostracodes dans les sédiments d’un ruisseau alpin (le Seebach, à Lunz, Autriche). – Verhandl. d. Int. Vereinig. f. theor. u. angew. Limnologie 22: 2053 - 2057 MCCANN, T. (Koord., 2008a): Carboniferous. – In: McCann, T. (Hrsg.): The Geology of Central Europe. Vol. 1 Precambrian and Palaeozoic. Geological Society London, S. 411 - 529 MCCANN, T. & KIERSNOWSKI, H. (Koord., 2008b): Permian. – In: McCann, T. (Hrsg.): The Geology of Central Europe. Vol. 1 Precambrian and Palaeozoic. Geological Society London, S. 530 - 597 MEISCH, C. (2009): Freshwater Ostracoda of Western and Central Europe. – In: Süßwasserfauna von Mitteleuropa 8/3, 522 S. RABAN, M., MERTMANN, D. & DOBMEIER, C.: GeoFeld. http://www.geo.fu-berlin.de/fb/e-learning/geofeld/ RADZINSKI, K.-H. (2008): Zechstein. – In: BACHMANN et al. (Hrsg.): Geologie von Sachsen-Anhalt, S. 160 - 178 ROTHE P. (2006): Die Geologie Deutschlands, 48 Landschaften im Portrait. – 2. Aufl., 240 S. SCHWAB, M. & EHLING, B.-C. (2008): Karbon. – In: Bachmann et al. (Hrsg.): Geologie von Sachsen-Anhalt, S. 110 - 140 SEIDEL, G. (1995): Geologie von Thüringen. – 2. Aufl., 556 S. SPANGENBERG, H.-J. (1973): Faunistisch-ökologische Untersuchungen an Gewässern von Gipshöhlen und im Grundwasser des Südharzes und Kyffhäusers. – Int. Revue Ges. Hydrobiol. 58 (4): 501 - 542 WAGNER, C.W. (1964): Ostracods as environmental indicators to Recent and Subrecent estuarine deposits of the Netherlands (with discussion). – In: Puri, S., Hrsg.: Ostracoda as ecological and palaeoecological indicators. Publicazioni della Stazione Zoologica di Napoli 33: 480 - 495 WICK, W. (1947): Aufbereitungsmethoden in der Mikropaläontologie. – 94. - 98. Jber. Naturhist. Ges. Hannover 1942 – 1947, 94 - 98: 35 - 41 WISSING, F.N. & HERRIG, E. (2001): Arbeitstechniken der Mikropaläontologie. – 191 S. WUNDERLICH, J. (2005): Abriss der Geologie des Kyffhäusers. – Geowiss. Exkf. u. Mitt. 225: 7 - 11 Anschrift der Autoren: B.Sc. Angewandte Geowissenschaften Anja Adler, anja.adler@student.uni-halle.de, und PD Dr. Dorothee Mertmann, dorothee.mertmann@geo.uni-halle.de, Fachgebiet Allgemeine Geologie, Institut für Geowissenschaften und Geographie, Martin Luther-Universität Halle-Wittenberg, Von Seckendorff-Platz 3, 06120 Halle Wir danken der Schriftleitung der Mitteilungen des Verbandes deutscher Höhlen- und Karstforscher für die freundliche Genehmigung, diesen Beitrag ebenfalls veröffentlichen zu dürfen. Weiterer Nachdruck oder Veröffentlichung bzw. Verbreitung in anderen elektronischen Medien nur mit schriftlicher Genehmigung der Schriftleitung. |